
ЁЁЁЁВЊЛљЖћ(I. H. Burkill)дкЁЖШЫЕФЯАЙпгыддХржВЮяЕФЦ№дДЁЗ(1951)жаЯЕЭГЕиПМжЄСЫжВЮяЫцШЫРрЪЯзхЕФЛюЖЏЁЂЯАЙпКЭЧЈсуЖјбБЛЏЕФЙ§ГЬЃЌТлжЄСЫЖЋАыЧђЖржжддХржВЮяЕФЦ№дДЃЌШЯЮЊЭпЮЌТхЗђЗНЗЈбЇЩЯжївЊШБЕуЪЧ“ШЋВПжЄОнЖМШЁзджВЮяЖјВЛЮЪддХржВЮяЕФШЫЁЃ”ЫћЬсГігАЯьбБЛЏКЭддХржВЮяЦ№дДЕФвЛаЉживЊЙлЕуЃЌШч“бБЛЏгЩздШЛВњЕигыаТВњЕижЎМфЕФВюБ№Жјв§Ц№ЁЃ”ЖдбБЛЏРДЫЕ“ИєРыЕФМлжЕЪЧОјЖдживЊЕФЁЃ”
ЁЁЁЁДяСжЖй(C. D. Darlington)РћгУЯИАћбЇЗНЗЈДгШОЩЋЬхЗжЮіддХржВЮяЕФЦ№дДЃЌВЂИљОнаэЖрШЫЕФвтМћЃЌНЋЪРНчддХржВЮяЕФЦ№дДжааФЛЎЮЊ9ИіДѓЧјКЭ4ИібЧЧјЃЌМД(1)ЮїФЯбЧжо;(2)ЕижаКЃЃЌИНХЗжобЧЧј;(3)АЃШћЖэБШбЧЃЌИНжаЗЧбЧЧј;(4)жабЧ;(5)гЁЖШ - УхЕщ;(6)ЖЋФЯбЧ;(7)жаЙњ;(8)ФЋЮїИчЃЌИНББУР(дкЭпЮЌТхЗђЛљДЁЩЯдіМгЕФвЛИіжааФ)МАжаУРбЧЧј;(9)УиТГЃЌИНжЧРћМААЭЮї - АЭРЙчбЧЧјЁЃЫћЕФЛЎЗжГ§СЫдіМгХЗжобЧЧјвдЭтЃЌЛљБОЩЯгыЭпЮЌТхЗђЕФЛЎЗжЯрНќЁЃ
ШуПЦЗђЫЙЛљЕФддХржВЮяДѓЛљвђжааФ
ЁЁЁЁШуПМЗђЫЙЛљ(Ї. ЇЎ. ЇЈЇхЇмЇрЇгЇуЇмЇкЇл)1970ФъЬсГіВЛЭЌзїЮяЮяжжЕФЕиРэЛљвђаЁжааФДя100грДІжЎЖрЃЌЫћШЯЮЊетжжаЁжааФЕФБфвьжжРрЖдзїЮяг§жжгаживЊЕФРћгУМлжЕЁЃЫћЛЙНЋЭпЮЌТхЗђШЗЖЈЕФ8ИіддХржВЮяЦ№дДжааФЫљАќРЈЕФЕиЧјЗЖЮЇМгвдРЉДѓЃЌВЂдіМгСЫ4ИіЦ№дДжааФЃЌЪЙжЎФмАќРЈЫљгавбЗЂЯжЕФддХржВЮяжжРрЁЃЫћГЦет12ИіЦ№дДжааФЮЊДѓЛљвђжааФЁЃет12 ИіДѓЛљвђжааФ(ЭМ1-2)ЁЃДѓЛљвђжааФЛђЖрбљЛЏБфвьЧјгђЖМАќРЈзїЮяЕФдЩњЦ№дДЕиКЭДЮЩњЦ№дДЕиЁЃ1979ФъКЩРМг§жжбЇМвдѓЮФ(A. C. Zeven)дкгыШуПМЗђЫЙЛљКЯБрЕФЁЖддХржВЮяМАЦфНќдЕжВЮяжааФДЧЕфЁЗжаЃЌАД12ИіЖрбљаджааФСаШы167ПЦ2 297жжддХржВЮяМАЦфНќдЕжВЮяЁЃЪщжаШЯЮЊдкДЫ12ИіЦ№дДжааФжаЃЌвдЖЋбЧ(жаЙњ - УхЕщ)ЁЂНќЖЋКЭжаУРШ§ЧјЪЧХЉвЕЕФвЁРКЃЌЖдддХржВЮяЕФЦ№дДЙБЯззюДѓЁЃШЛЖјЃЌгЩгк12Иі“жааФ”ИВИЧЕФЗЖЮЇЙ§гкЙуЗКЃЌМИКѕАќРЈЕиЧђЩЯГ§СНМЋвдЭтЕФШЋВПТНЕиЁЃ
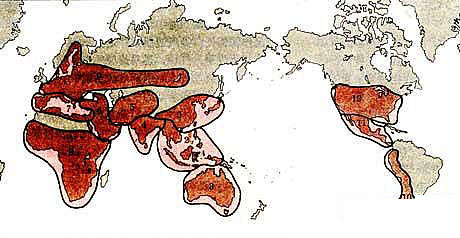
ЁЁЁЁЭМ1-2ЁЁШуПЦЗђЫЙЛљЕФддХржВЮяДѓЛљвђжааФЁЃ(1)жаЙњ - ШеБОжааФЃЌжаЙњЮЊГѕЩњЛљвђжааФЃЌШеБОЮЊДЮЩњЛљвђжааФ;(2)ЖЋФЯбЧжааФЃЌгЁЖШжЇФЧЁЂгЁЖШФсЮїбЧКЭТэРДШКЕКЕШ;(3)АФДѓРћбЧжааФ;(4)гЁЖШжааФ;(5)жабЧжааФЃЌАќРЈАЂИЛКЙЁЂЫўМЊПЫКЭЮкзШБ№ПЫ(ЬьЩНЮїВП)ЕШ;(6)ЮїбЧжааФЃЌАќРЈЭСПтТќ;(7)ЕижаКЃжааФ;(8)ЗЧжожааФ;(9)ХЗжо - ЮїВЎРћбЧжааФЃЌ(10)ФЯУРжааФ;(11)жаУРКЭФЋЮїИчжааФ;(12)ББУРжааФ(в§здЁЖжаЙњДѓАйПЦШЋЪщ·ХЉвЕIIЁЗ)ЁЃ
ЁЁЁЁЙўРМ(J. R. HarlanЃЌ1971)ШЯЮЊЃЌдкЪРНчЩЯФГаЉЕиЧј(ШчжаЖЋЁЂжаЙњББВПКЭжаУРЕиЧј)ЗЂЩњЕФбБЛЏгыЭпЮЌТхЗђЦ№дДжааФФЃЪНЯрЗћЃЌЖјдкСэвЛаЉЕиЧј(ШчЗЧжоЁЂЖЋФЯбЧКЭФЯУР—ЖЋгЁЖШШКЕК)ЗЂЩњЕФбБЛЏдђгыЦ№дДжааФФЃЪНВЛЗћЁЃЫћИљОнзїЮябБЛЏжаРЉЩЂЕФЬиЕуЃЌАбддХржВЮяЗжЮЊ5РрЁЃ
ЁЁЁЁ(1)ЭСЩњаЭЁЁжВЮядквЛИіЕиЧјбБЛЏКѓЃЌДгЮДРЉЩЂЕНЦфЫћЕиЧјЁЃШчЗЧжоЕОЁЂАЃШћЖэБШбЧАХНЖЕШЯЪЮЊШЫжЊЕФзїЮяЁЃ
ЁЁЁЁ(2)АыЭСЩњаЭЁЁБЛбБЛЏЕФжВЮяжЛдкСкНќЕиЧјРЉЩЂЃЌШчдЦФЯЩНщЋЁЂЮїВиЙтКЫЬвЕШЁЃ
ЁЁЁЁ(3)ЕЅвЛжааФЁЁдкдВњЕибБЛЏКѓбИЫйДЋВЅЕНЙуДѓЕиЧјЃЌУЛгаДЮЩњжааФЃЌШчЯ№НКЁЂПЇЗШЁЂПЩПЩЁЃ
ЁЁЁЁ(4)гаДЮЩњжааФЁЁзїЮяДгвЛИіУїШЗЕФГѕЩњЦ№дДжааФж№НЅЯђЭтРЉЩЂЃЌдквЛИіЛђМИИіЕиЧјаЮГЩДЮЩњЦ№дДжааФЃЌШчЦЯЬбЁЂЬвЁЃ
ЁЁЁЁ(5)ЮожааФЁЁУЛгаУїШЗЕФЦ№дДжааФЃЌШчЯуНЖЁЃ
ЁЁЁЁЭЦМідФЖСЃК
